细胞记忆机制有助于在间歇性接触合成代谢类固醇后长时间内实现肌肉超负荷肥大概括介绍
之前进行的力量训练(无论是否使用合成代谢类固醇)有助于肌肉质量的恢复,即使在长时间不活动之后也是如此。基于体内和体外显微镜,我们在此提出一种驻留在肌肉细胞中的细胞记忆机制。雌性小鼠接受丙酸睾酮治疗 14 天,肌核数量增加 66%,纤维横截面积增加 77%。停药三周后,纤维尺寸减小到与假治疗动物相同的水平,但细胞核数量至少在 3 个月内保持升高(> 小鼠寿命的 10%)。此时,当富含肌核的肌肉暴露于超负荷运动 6 天时,纤维横截面积增加了 31%,而对照肌肉没有显着增长。我们认为,持久的肌核数量增加构成了一种细胞记忆,促进了随后的肌肉超负荷肥大。我们的发现可能会对兴奋剂违规者的禁赛时间产生影响。由于老年人产生新肌核的能力受损,我们的数据也引发了人们的猜测,即年轻时进行力量训练可能对衰老有益。
关键点
训练室的民间传说表明,之前的力量训练(无论是否使用合成代谢类固醇)有助于在长时间不活动后重新获得肌肉质量。这种“肌肉记忆”以前被归因于运动学习,但我们的数据表明,肌肉纤维本身中存在细胞记忆。
肌纤维有多个细胞核,肌肉质量增加时细胞核的数量也会增加。
当小鼠短暂接受类固醇治疗时,肌肉质量和细胞核数量增加。随后停药 3 个月,肌肉质量恢复正常,但过量的细胞核仍然存在。当这些肌肉承受超负荷时,它们在 6 天内增长了 30%,而对照组的增长则微不足道。
我们的数据表明,之前的力量训练可能会对以后的生活有益,而短暂接触合成代谢类固醇可能会产生持久的提高运动表现的效果。
介绍
肌纤维与神经元一起构成了身体两种主要有丝分裂后组织之一。按体积计算,肌纤维是哺乳动物中最大的细胞(Bruusgaard等人, 2003 年),是脊椎动物中极少数合胞体之一。永久纤维可包含数百个细胞核(Bruusgaard等人, 2003 年),每个细胞核都被自己的合成机制所包围,为其附近的局部区域合成蛋白质(Hall 和 Ralston,1989 年;Pavlath等人, 1989 年)。在超负荷肥大期间,纤维体积的增加之前会掺入来自间质肌肉干细胞(卫星细胞;Schiaffino等人, 1976 年;Bruusgaard等人, 2010 年;Wang 和 Rudnicki,2012 年)的新肌核。最近的证据表明,即使在随后的肌肉质量损失的情况下,这些“额外”的细胞核也是永久性的,或者至少是长期存在的(Wada等人, 2002 年;Aravamudan等人, 2006 年;Bruusgaard 和 Gundersen,2008 年;Gundersen 和 Bruusgaard,2008年;Bruusgaard等人, 2010 年、2012 年;van der Meer等人, 2011 年)。因此,虽然肌肉肥大在肌肉质量方面是完全可逆的,但先前的肥大情况似乎会以肌核数量增加的形式在肌肉纤维上留下持久的印记。
我们在此报告,使用类固醇进行间歇性治疗可诱导更大的纤维和更多的肌核。即使在停药后肌肉大小恢复正常数月后,这些额外的肌核也不会消失。然而,当这些肌肉受到超负荷锻炼时,它们的生长速度比对照组快得多。
方法
动物实验
实验采用体重 25-30 克的雌性 NMRI 小鼠。动物实验经挪威动物研究局批准,并根据 1974 年 12 月 20 日的《挪威动物福利法》进行。挪威动物研究局提供管理,以确保设施和实验符合该法案、1996 年 1 月 15 日的国家法规以及 1986 年 3 月 18 日的《欧洲实验和其他科学目的脊椎动物保护公约》。
所有非终点实验均采用含 2% 异氟烷的吸入气体麻醉。终点实验采用腹膜内注射 5 μl (g 体重) −1 Equithesin (Ullevål Sykehus,挪威;42.5 mg ml −1水合氯醛和 9.7 mg ml −1戊巴比妥)。所有成像和手术均在深度麻醉下进行。通过捏肢体的跖骨区域定期检查麻醉深度,如有必要,可给予额外剂量。动物在深度麻醉时通过颈部脱臼处死。
为了增加趾长伸肌 (EDL) 的负荷,切除了胫骨前肌远端的约三分之二。通过切除腓肠肌和跖肌的类似部分来增加比目鱼肌的负荷。
通过在颈部皮肤上开一个 5 毫米长的切口,将含有丙酸睾酮的颗粒或假颗粒(美国创新研究公司 A-211,2.5 毫克,释放时间为 21 天)植入皮下。
血液中睾酮水平分析
在六只小鼠中,在植入颗粒前、植入后 1 天和 14 天以及取出颗粒后 7 天抽取血液样本(见图2A中的插图)。分析在奥斯陆大学医院经批准的(NS-EN ISO/IEC 17025)激素实验室进行,使用来自 Orion Diagnostica(芬兰埃斯波)的放射免疫分析 (RIA) 试剂盒,经乙醚提取后进行。
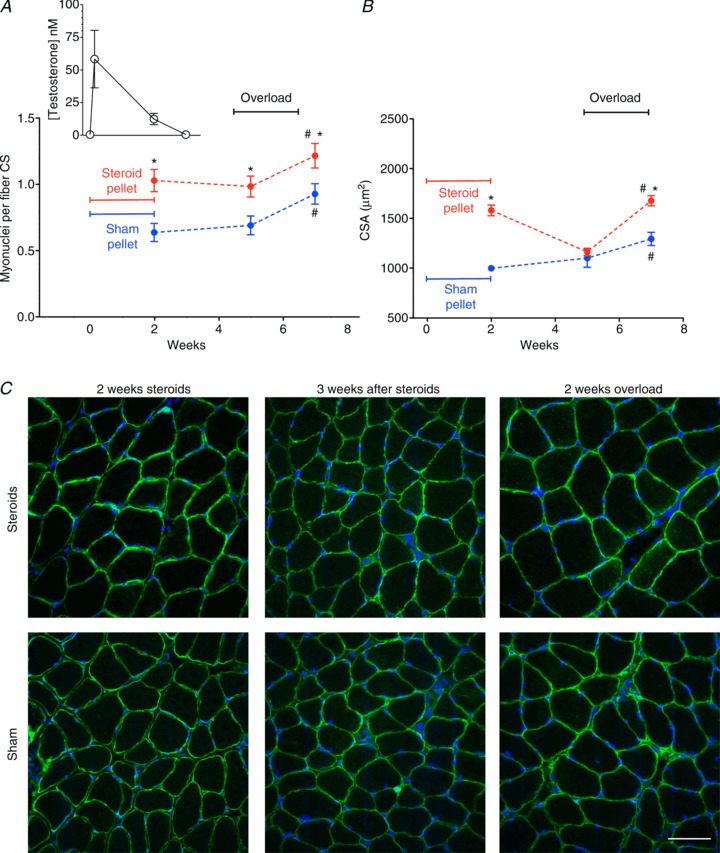
图 2. 先前服用合成代谢类固醇对横截面积 (CS; A )上每根纤维的肌核数量和纤维横截面积 (CSA;B ) 的影响。
A中的插图显示血液睾酮浓度,与主图位于同一时间轴上。 *与假手术有显著差异(P < 0.05);#与超负荷有显著差异(P < 0.05)。每个数据点代表来自 6 块肌肉的 300 个纤维。C为使用合成代谢类固醇或假手术颗粒治疗 2 周、去除颗粒 3 周后以及随后进行 2 周功能超负荷后,用 Hoechst 染料 33342(蓝色)染色以标记 DNA 和抗肌萎缩蛋白抗体(绿色)的冷冻切片的代表性显微照片。比例尺为 50 μm。
体内肌核成像
为了在体内标记肌核,向 EDL 中的单根纤维注射一种溶液,该溶液含有溶解在注射缓冲液(10 m m NaCl、10 m m Tris,pH 7.5、0.1 m m EDTA 和 100 m m葡萄糖酸钾)中的 5′ – TRITC 标记的随机 17 聚体寡核苷酸,该寡核苷酸具有硫代磷酸酯化骨架(Yorkshire Biosciences Ltd,英国赫斯灵顿)。寡核苷酸显然通过主动运输被吸收到注射纤维内的细胞核中,在我们的实验中仅用作活体核染料。该方法此前已详细描述(Utvik等人, 1999 年;Bruusgaard等人, 2003 年、2010 年)。在本研究中,表面纤维被去除以露出肌肉内部的纤维。这样,我们就可以注射更能代表整个肌肉的纤维,而不是像以前那样只注射表面纤维。因此,材料中既包括表面纤维,也包括深层纤维。寡核苷酸包含随机选择的序列 TAGTCCTAAGTGGACGC,BLAST 分析证实,该序列在小鼠基因组中既没有以正向也没有以反向出现。
体内成像基本按照之前描述的方式进行(Balice-Gordon 和 Lichtman,1994 年;Utvik等人, 1999 年;Bruusgaard等人, 2003 年、2010 年)。通过在配备×20 0.3 NA 长工作距离水浸物镜的 Olympus BX-50WI 复式显微镜上获取相距 5 μm 的不同焦平面中的图像来分析 250–1000 μm 的纤维段。所有图像均使用 Andor iXion+ 相机获取,并由 Andor SOLIS 软件控制。通过将图像导入运行 Adobe Photoshop 和 NIH ImageJ 软件的 Macintosh 计算机,生成一个堆栈并用于计算段中的所有细胞核。通过评估每个堆栈中的所有图像来计算细胞核。插图图像(图 3C )由几个焦平面合成,以显示每个纤维段中的所有肌核。
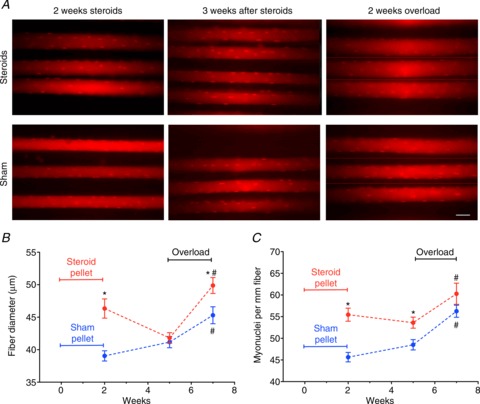
图 3.体内成像表明,先前使用合成代谢类固醇会增加每毫米纤维的肌核数量(A和B),并增强 EDL 肌肉对超负荷的后续肥大反应(A和C)。
* 与假手术有显著差异(P < 0.001);# 与超负荷有显著差异(P < 0.001)。每个数据点代表来自 6 至 8 块肌肉的 33 至 50 根纤维。C ,代表性体内肌肉图像,每张图像显示三根注射寡核苷酸的肌肉纤维标记肌核,以及在用合成代谢类固醇或假手术颗粒治疗 2 周后、在去除颗粒 3 周后以及随后进行 2 周的功能超负荷后的单个纤维的轮廓。在A中,发丝表示描绘不同焦平面时的边界。比例尺为 50 μm。
免疫组织化学
切除肌肉并嵌入 OCT Tissue-Tek 中,然后在融化的异戊烷中冷冻、轻微拉伸,并在−80°C 下储存,然后在 10 μm 处进行冷冻切片。随后将切片放入 1% 牛血清白蛋白中封闭,并用抗肌萎缩蛋白单克隆抗体(Abcam,克隆 MANDYS8 ab7163)染色。二抗是山羊抗鼠 TRITC 结合 IgG(Sigma T7782)。两种抗体均以 1:200 的稀释度使用。最后使用 Hoechst 染料 33342(Invitrogen;PBS 中0.1 μg ml −1 )对细胞核进行共染色。如前所述(Gundersen & Bruusgaard,2008;Bruusgaard等人, 2012),为了确保仅将肌纤维内的细胞核纳入分析,肌核被定义为几何中心位于肌营养不良蛋白环内缘内的细胞核。
肌球蛋白重链纤维类型的组织学分析按照之前描述的方法进行(Lunde等人, 2011 年),使用以下单克隆抗体组:BA-D5 (I)、SC-71 (IIa)、6H1 (IIX) 和 BF-F3 (IIb)。所有这些都是从 ATCC/LGC 标准中获得的杂交瘤原种内部培养的。为了估计纤维类型组成,在整个肌肉部分上放置了一个网格,并确定位于网格交叉点的每根纤维的纤维类型。
统计数据
通过方差分析 (ANOVA) 和 Bonferroni 多重比较校正来检验差异。数值以平均值 ± SEM 给出。
结果
我们研究了合成代谢类固醇和超负荷运动对体内肌肉细胞的影响。成年雌性小鼠皮下植入释放丙酸睾酮的颗粒,持续 14 天。两组中的一些动物还切除了协同肌,以便在药物治疗期间使氧化/缓慢的比目鱼肌或糖酵解/快速的伸趾长肌 (EDL) 超负荷。
在比目鱼肌中,与假颗粒治疗相比,单独使用类固醇治疗可使肌核数量增加 66%,而单独使用超负荷治疗可使肌核数量增加 51%(图 1 A)。当类固醇和超负荷相结合时,增幅达到 92%(图 1 A)。在 EDL 中观察到了定性相似但不那么明显的效果(图 1 B)。类固醇的使用也已被证明可以增加人类的肌核数量(Eriksson等人, 2005 年)。
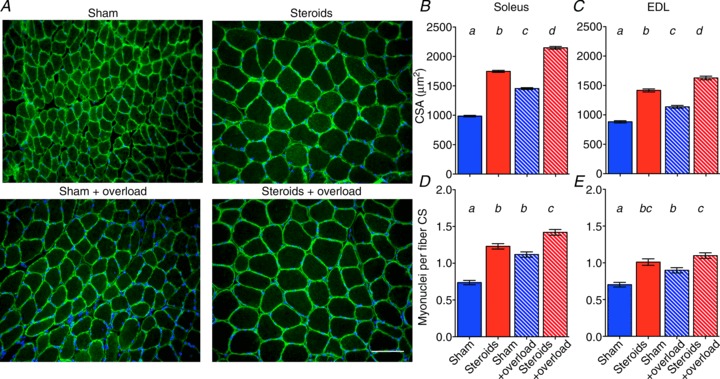
图 1. 合成代谢类固醇和功能超负荷对横截面积 (CSA) 和每根纤维横截面上的肌核数量 (CS) 的影响。
A,用 Hoechst 染料 33342(蓝色)染色的冷冻切片的代表性显微照片,以标记 DNA,以及抗肌萎缩蛋白的抗体(绿色),这些肌肉来自经过 2 周的假治疗或类固醇治疗,同时有或没有同时进行功能超负荷的比目鱼肌。比例尺为 100 微米。比目鱼肌(B和D)和 EDL(C和E)的 CSA 和每个纤维横截面(CS)的肌核数量。条形图显示平均值 ± SEM。列上方的字母a–d表示统计学上显着差异( P < 0.01);标有相同字母的列在统计学上没有差异。每列代表来自 6 块肌肉的 317–896 条纤维。
虽然类固醇对肌肉纤维类型没有影响(补充图 S1),但纤维大小的变化大致反映了肌核的变化(图 1C和E )。在比目鱼肌中,单独使用类固醇治疗与假颗粒治疗相比,纤维横截面积 (CSA) 增加了 77%,而单独超负荷只诱导了 48% 的肥大。当类固醇和超负荷结合在一起时,观察到 118% 的肥大,因此两种治疗的效果大致是累加的。至于肌核,在 EDL 中观察到质量相似但不那么明显的效果(图 1D )。这些在小鼠中发现的效果与阻力运动和睾酮治疗在人类中的效果相似(Bhasin等人, 1996 年)。
我们接下来要问的是,类固醇是否会对肌核数量产生长期影响,以及在停药后,肌核大小恢复正常,额外的肌核是否有助于肌肉质量的增加。图2 A(插图)显示了植入丸剂期间和取出丸剂后血液睾酮浓度的变化,我们确定在取出丸剂后不到一周,睾酮水平就低于检测限(<0.05 n m)。在取出丸剂 3 周后检查比目鱼肌时,肌核数量并没有显著减少,平均比假手术组高 42%(图 2 A)。然而,CSA 完全逆转,类固醇和假手术治疗肌肉的纤维大小难以区分(图 2 B和C)。当此时以及随后的 14 天内引入超负荷时,类固醇组表现出 44% 的肥大,而假手术治疗小鼠的细胞仅生长了 17% (图 2B和C )。
尽管 EDL 对类固醇的反应不如比目鱼肌强烈(图 1),但进一步研究 EDL 仍然很重要。比目鱼肌是一种氧化姿势性肌肉,可能与力量表现不太相关。此外,EDL 更适合体内成像,可以直接在活体动物的单纤维中原位观察肌核。这是可取的,因为传统的离体组织学技术会在这种类型的实验中引入混淆解释的伪影(Gundersen 和 Bruusgaard,2008 年;Bruusgaard等人, 2012 年)。EDL中的体内单纤维观察支持了比目鱼肌的离体分析。因此,2 周的类固醇治疗导致肌节长度为 3 μm 时每毫米纤维长度的肌核数量从 46 个增加到 56 个,即增加了 20%(图 3 A和C)。纤维直径增加了 18%,假设纤维为圆柱形,则 CSA 增加了 39%(图 3B和C )。去除颗粒三周后,类固醇组和假手术组的纤维直径难以区分(图 3B和C ),而任何组的细胞核数量均未发生显著变化(图 3A和C )。此时引入超负荷时,纤维直径的增加表明,在 14 天内,类固醇组的 CSA 增长了 42%,假手术组的 CSA 增长了 21%(图 3B和C )。
为了证明长期记忆,我们还研究了在去除颗粒 3 个月后间歇性类固醇治疗的效果。圈养的 NMRI 小鼠可活约 2 年(Lhrke等人, 1984 年),因此 3 个月约占其寿命的 12%,相当于人类的 10 年。此时,虽然 CSA 或纤维类型没有差异(补充图 S2),但类固醇组的细胞核数量比假手术组高 28%(图 4 A)。当引入超负荷时,类固醇组的 CSA 在前 6 天增加了 31%,而假手术组仅增加了 6%(不显著;图 4 B和C)。此后两组平行增长,但超负荷 14 天后类固醇组的 CSA 仍高 20%(图4 B)。
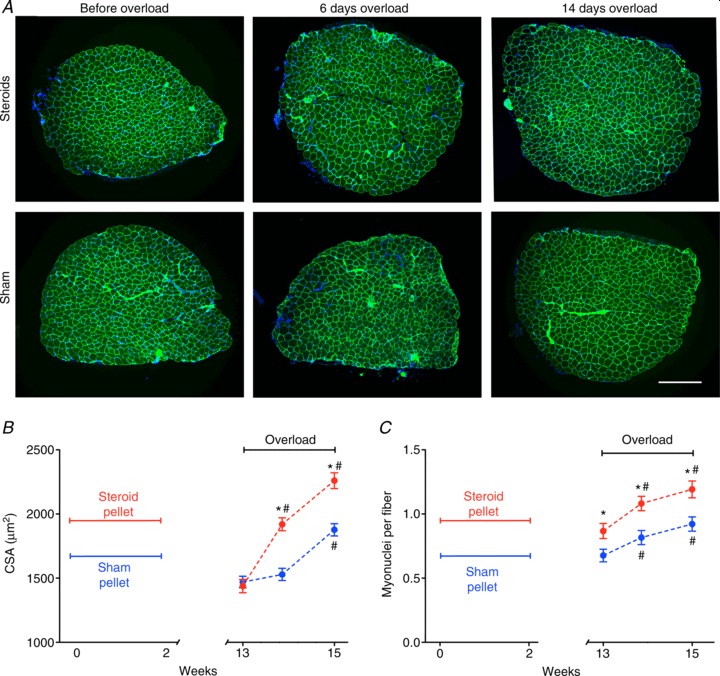
图 4. 短期使用合成代谢类固醇对肌核(A)和 EDL 肌肉因超负荷而引起的纤维生长的长期影响(B和C)。
*与假手术有显著差异(P < 0.001);#与超负荷有显著差异(P < 0.001)。每个数据点代表 6 块肌肉的 669-856 条纤维。C ,在使用合成代谢类固醇或假手术颗粒治疗 2 周后立即用 Hoechst 染料 33342(蓝色)染色以标记 DNA 和抗肌萎缩蛋白抗体(绿色)的冷冻切片的代表性显微照片,以及 3 个月后肌肉功能超负荷 6 天和 14 天的显微照片。比例尺为 300 μm。
讨论
我们在此为一种新的表观遗传机制提供了证据,即肌肉合体中细胞核数量增加有助于恢复先前较大的肌肉质量。这种机制可能具有进化重要性,因为它可以让例如执行季节性力量要求高的任务的个体快速恢复力量,而不必保持较大的永久肌肉质量。在现代社会中,人们普遍观察到,先前的力量训练似乎使人们在以后的生活中更容易恢复肌肉质量,即使在长时间不活动和质量损失之后也是如此,这种现象被称为“肌肉记忆”(Staron等人, 1991 年;Taaffe 和 Marcus,1997 年)。此前尚无已知的肌肉细胞记忆机制,先前训练的长期影响完全归因于中枢神经系统的运动学习(Rutherford 和 Jones,1986 年)。
肌肉肥大和记忆的机制
在没有完善的细胞内运输系统的情况下,肌细胞仍能维持跨越广阔细胞内距离的较大细胞质体积的能力似乎与整个细胞中具有大量最佳分布的细胞核有关(Bruusgaard等人, 2003 年;Metzger等人, 2012 年;Qaisar等人, 2012 年)。在许多肌病中通常观察到细胞核错位,例如,在 Emery-Dreifuss 综合征中,肌营养不良症通常与核膜基因突变有关(Puckelwartz 和 McNally,2011 年)。在超负荷期间,从卫星细胞中募集新细胞核先于细胞质体积增加(Bruusgaard等人, 2010 年),但之前的文献得出结论,肌核在随后的萎缩过程中丢失,表明细胞结构完全可逆。然而,这些发现主要基于传统的组织学,可能无法提供肌核和组织中其他核(例如卫星细胞)之间的明确分离。然而,最近(以及在本文中),单纤维的体内和离体成像显示,没有肌核丢失,并且在萎缩期间“额外”的核仍留在细胞中(Wada等人2002 年;Aravamudan等人2006 年;Bruusgaard 和 Gundersen,2008 年;Gundersen 和 Bruusgaard,2008 年;Bruusgaard等人2010 年、2012 年;Jackson等人2012 年a期)。这代表了细胞结构滞后,使我们能够假设核数量的增加可能是与肌肉质量相关的肌肉记忆的一种机制。目前的数据表明,存在这样一种有益的细胞记忆,它存在于肌肉细胞本身中。
在正常动物中,新细胞核的添加似乎伴随着各种条件下的肥大生长(Enesco & Puddy, 1964 ; Moss, 1968 ; Cheek et al. 1971 ; Schiaffino et al. 1976 ; Seiden, 1976 ; Cabric & James, 1983 ; Cabric et al. 1987 ; Giddings & Gonyea, 1992;Allen等,1999;Bruusgaard等, 2012 。据推测,肌核的增加主要与可能诱发肌肉损伤的情况有关,例如协同肌肉消除后,或剧烈运动伴有离心收缩后(Schiaffino等人, 2013 年),这意味着它与肌肉修复的关系比与现有纤维的肥大性生长更密切。然而,肌核的增加似乎也伴随着与睾酮(本数据和Kadi,2008 年)或 IGF-1(Barton-Davis等人, 1999 年)等激素治疗相关的肥大,这些激素不太可能造成损伤。
在超负荷肥大研究中, 3 H-胸苷标记(Aloisi等人, 1973 年)和体内成像(Bruusgaard等人, 2010 年)均表明,在 CSA 发生任何明显变化之前,新的肌核就已经被整合,这表明肌核在肥大中起着因果作用。然而,我们并不认为肌核数量是决定肌纤维大小的唯一因素;例如,我们在此表明,尽管类固醇治疗诱导的肌核数量较多,但肌纤维会收缩,当类固醇停用时,肌纤维并不比具有较少核的对照纤维大。此外,在后肢悬吊后纤维重新长回正常大小的过程中,没有增加细胞核(Bruusgaard等人, 2012 年;Jackson等人, 2012 b),这使我们认为细胞核的数量可能代表纤维在其历史上的最大尺寸,而不是其当前尺寸(Bruusgaard等人, 2012 年)。此外,使用与本文中使用的类似的体内成像方法,我们之前发现,相同大小的慢纤维中的细胞核数量多于快纤维中的细胞核数量(Bruusgaard等人, 2003 年)。这可能与慢肌的更高周转和对蛋白质合成的更高要求有关(Goldberg,1967 年;Kelly等人, 1984 年)。通常,在蛋白质周转率相对较高的情况下,细胞核数量和纤维大小之间似乎存在更强的相关性(Bruusgaard等人, 2006 年)。我们认为纤维大小取决于:(i)蛋白水解,(ii)每个细胞核的蛋白质合成和(iii)细胞核的数量。
多种基因操控小鼠表现出肥大但细胞核数量并未相应增加,例如过度表达蛋白质 Akt(Blaauw等人, 2009 年)、JunB(Raffaello等人, 2010 年)或 Ski(Bruusgaard等人, 2005 年)的小鼠,或携带肌生长抑制素基因功能丧失突变的小鼠(Amthor等人, 2009 年)。Ski 小鼠的收缩物质含量减少(Bruusgaard等人, 2005 年),肌生长抑制素小鼠的比力降低(Amthor等人, 2007 年;Mendias等人, 2011 年),这些异常可能与细胞核数量低有关。相反,Akt 小鼠在 3 周后似乎具有正常的比力(Blaauw等人, 2009 年)。类似地,在使用可诱导的 Pax7-白喉毒素 A 转基因消融 90% 的卫星细胞的小鼠中,经过 2 周的超负荷肥大后,特定力量似乎恢复正常(McCarthy等人, 2011 年)。
对于后一种模型,需要进行更多研究来确定没有正常增加肌核的肥大是否会持续一段时间,成本是否更高,或导致功能特性不理想(短期比力除外)。或者,没有增加细胞核的肥大可能代表机械冗余,这在生物系统中很常见。虽然我们不能推断其他实验模型或排除睾酮对肌肉有其他长期影响的可能性,而不是增加细胞核的数量,但我们认为,细胞核较多的肌肉似乎更适应快速再生,因此是肌肉记忆的承载者。
运动与公共健康中的肌肉记忆
老年动物 ( Carson等人, 1995 年;Alway等人, 2002 年)中,超负荷引起的肌肥大大大减弱,而人类中目前超过 50% 的个体在年龄 >80 岁时符合虚弱的临床标准 ( Matthews等人, 2011 年),这给西方老龄化人口带来了严重的健康问题 ( Dutta 和 Hadley,1995 年;Hughes 和 Schiaffino,1999 年)。然而,事实证明,即使在多年停止训练之后,老年人进行高强度阻力训练仍然有益,特别是如果个体保持中等强度的训练计划 ( Smith等人, 2003 年)。然而,产生新肌核的能力会随着年龄的增长而受损 ( Schultz 和 Lipton,1982 年),可能是因为 Notch 信号减弱 ( Conboy等人, 2003 年;Conboy 和 Rando,2005 年)。我们的数据表明存在与肌核相关的长期肌肉记忆。需要进行调查以确定人类是否存在与力量训练相关的类似核相关肌肉记忆;如果存在,则应考虑针对年轻人进行力量训练的公共卫生建议,因为它可能有助于在衰老过程中更容易保持肌肉质量。
我们的数据表明,至少在小鼠中,一次睾酮使用可以募集一个持久的过量肌核池,并且在没有进一步使用类固醇的情况下,通过阻力训练恢复肌肉质量的能力持续增强。因此,即使是偶尔滥用药物,对运动员来说,其益处也可能是持久的,即使不是永久的。我们的数据表明,应该重新考虑《世界反兴奋剂条例》中关于类固醇使用定罪后仅 2 年禁赛的规定(WADA,2009 年)。
致谢
我们感谢 Siobhan L. Anton 对手稿的评论。
感谢PED大神的转载普及,任何问题咨询可以联系微信:PEDmaster001
